Mitochondrial DNA (mtDNA) contains instructions for building important cellular machines. We have populations of mtDNA inside each of our cells -- almost like a population of animals in an ecosystem. Indeed, mitochondria were originally independent organisms, that billions of years ago were engulfed by our ancestor's cells and survived -- so the picture of mtDNA as a population of critters living inside our cells has evolutionary precedent! MtDNA molecules replicate and degrade in our cells in response to signals passed back and forth between mitochondria and the nucleus (the cell's "control tower"). Describing the behaviour of these population given the random, noisy environment of the cell, the fact that cells divide, and the complicated nuclear signals governing mtDNA populations, is challenging. At the same time, experiments looking in detail at mtDNA inside cells are difficult -- so predictive theoretical descriptions of these populations are highly valuable.
Why should we care about these cellular populations? MtDNA can become mutated, wrecking the instructions for building machines. If a high enough proportion of mtDNAs in a cell are mutated, our cells struggle and we get diseases. It only takes a few cells exceeding this "threshold" to cause problems -- so understanding the cell-to-cell distribution of mtDNA is medically important (as well as biologically fascinating). Simple mathematical approaches typically describe only average behaviours -- we need to describe the variability in mtDNA populations too. And for that, we need to account for the random effects that influence them.
Why should we care about these cellular populations? MtDNA can become mutated, wrecking the instructions for building machines. If a high enough proportion of mtDNAs in a cell are mutated, our cells struggle and we get diseases. It only takes a few cells exceeding this "threshold" to cause problems -- so understanding the cell-to-cell distribution of mtDNA is medically important (as well as biologically fascinating). Simple mathematical approaches typically describe only average behaviours -- we need to describe the variability in mtDNA populations too. And for that, we need to account for the random effects that influence them.
 |
| In our cells, signals from the "control tower" nucleus lead to the replication (orange) and degradation (purple) of mtDNA. These processes affect mtDNA populations that may contain normal (blue) and mutant (red) molecules. Our mathematical approach -- extending work addressing a similar but simpler system -- describes how the total number of machines, and the proportion of mutants, is likely to behave and change with time and as cells divide. |
In the past, we have used a branch of maths called stochastic processes to answer questions about the random behaviour of mtDNA populations. But these previous approaches cannot account for the "control tower" -- the nucleus' control of mtDNA. To address this, we've developed a mathematical tradeoff -- we make a particular assumption (which we show not to be unreasonable) and in exchange are able to derive a wealth of results about mtDNA behaviour under all sorts of different nuclear control signals. Technically, we use a rather magical-sounding tool called "Van Kampen's system size expansion" to approximate mtDNA behaviour, then explore how the resulting equations behave as time progresses and cells divide.
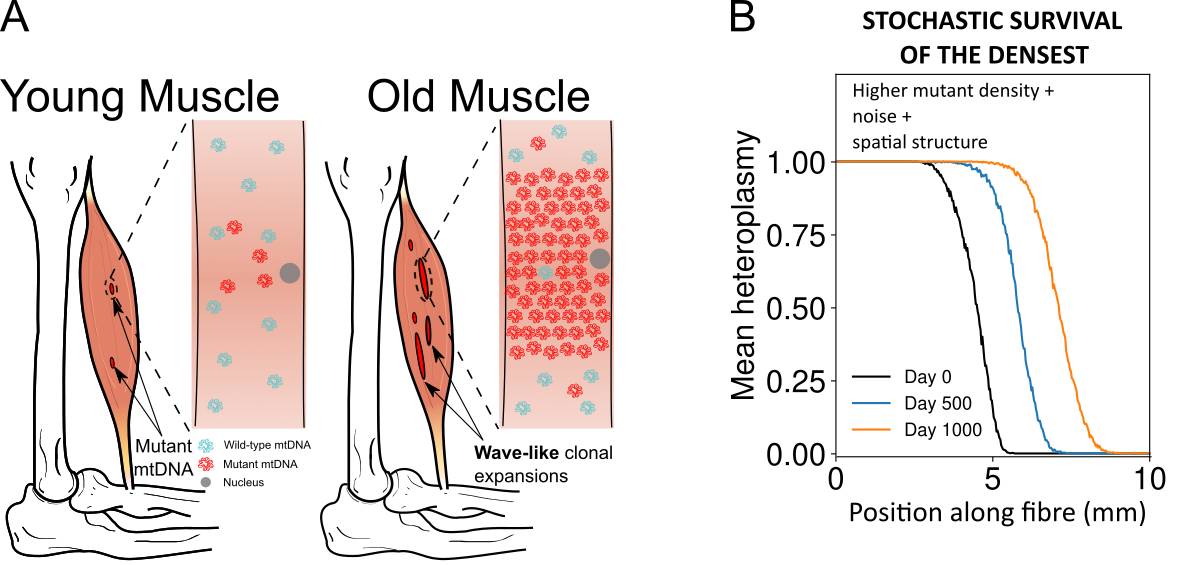
Our approach shows that the cell-to-cell variability in heteroplasmy (the potentially damaging proportion of mutants in a cell) generally increases with time, and surprisingly does so in the same way regardless of how the control tower signals the population. We're able to update a decades-old and commonly-used expression (often called the Wright formula) for describing heteroplasmy variance, so that the formula, instead of being rather abstract and hard to interpret, is directly linked to real biological quantities. We also show that control tower attempts to decrease mutant mtDNA can induce more variability in the remaining "normal" mtDNA population. We link these and other results to biological applications, and show that our approach unifies and generalises many previous models and treatments of mtDNA -- providing a consistent and powerful theoretical platform with which to understand cellular mtDNA populations. The article is in the American Journal of Human Genetics here and a preprint version can be viewed here. Crossed from here.